La célula eucariota se distingue también por una red proteica, el citoesqueleto, que sitúa y desplaza los organelos en la célula. Todos los vegetales, los hongos y los animales son eucariotas. Su DNA está en uno de los organelos, el núcleo.
Además del retículo endoplásmico y el complejo de Golgi, que intervienen en la síntesis de ciertas proteínas, los otros dos compartimientos que desempeñan un papel sustancial ya que suministran energía a la célula son las mitocondrias (del griego mitos, filamento, y khondros, grano), las cuales se ocupan de la respiración celular, y los plastos de los vegetales, donde tiene lugar la fotosíntesis o conversión de la energía luminosa en energía química.
Algunos organelos pueden desaparecer y volverse a formar. Es el caso del núcleo durante la división celular, del aparato de Golgi y del retículo. Pero las mitocondrias y los plastos proceden siempre de la división de mitocondrias y plastos preexistentes. A finales del siglo XIX, esta particularidad hacía pensar ya en bacterias que se dividían por bipartición en el citoplasma. Las mitocondrias y los plastos eran organismos vivos, instalados en simbiosis en la célula.
En 1883, el investigador alemán A. Schimper observó: “Si se establece definitivamente que los plastos nunca se forman de novo en la célula huevo, entonces su situación en la célula donde se encuentran recuerda la de los simbiontes. Es posible que una planta verde no sea otra cosa que la unión entre un organismo incoloro y un microbio que posee los pigmentos clorofílicos.” Otros autores le pisaron los talones a principios del siglo XX, pero toparon, lamentablemente, con la incredulidad de sus colegas. Ocurrió como con la deriva de los continentes de Alfred Wegener: los argumentos propuestos no bastaron para afianzar la intuición.
En particular, fracasaron todos los intentos de aislamiento y cultivo in vitro del microbio verde. Varios autores propusieron para estos organelos un origen autógeno a partir de repliegues de la membrana plasmática. Los progresos de la biología iban a brindar otros argumentos a la idea según la cual las mitocondrias y los plastos derivan de bacterias.
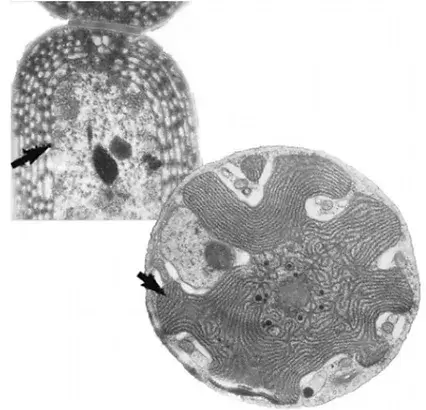
La microscopia electrónica demostró que los organelos de los eucariotas están limitados por una membrana lipídica simple, salvo tres de ellos: el núcleo, rodeado por membranas provistas de poros que lo ponen en continuidad con el citoplasma, y los plastos y las mitocondrias, separados del citoplasma por dos membranas continuas. Además, la organización de la estructura de estos dos últimos organelos evoca la de ciertas bacterias. Por ejemplo, las membranas fotosintéticas o tilacoides de los plastos de ciertas algas (las algas rojas) evocan en gran medida a los tilacoides de bacterias fotosintéticas, las cianobacterias. Los adelantos de la bioquímica pusieron de manifiesto otros parecidos sorprendentes. Los lípidos de las membranas que rodean a las mitocondrias existen en ciertas bacterias, pero no en otras partes de la célula eucariota. En cuanto a los lípidos característicos de las membranas de los plastos, se les encuentra en las cianobacterias. Además, las mitocondrias y los plastos desempeñan ciertas funciones metabólicas que efectúan también ciertos procariotas libres.
La fotosíntesis se consuma de manera casi idéntica en las cianobacterias y los plastos. Las reacciones de oxidorreducción ligadas a la actividad energética de las mitocondrias y de los plastos requieren la intervención de cadenas de transferencia de electrones membranosas parecidas a las de los procariotas: colaboran en ellas unas complejas proteínas homólogas, como los citocromos.
Pero el argumento principal fue el descubrimiento de un genoma en los plastos en 1962 y luego en las mitocondrias al año siguiente. Este DNA se parece al de los procariotas:
no está aislado dentro del organelo y se presenta en forma de varias copias idénticas de una molécula circular. El DNA se duplica y se transfiere durante la bipartición del organelo como en el caso de los procariotas libres. En las mitocondrias y los plastos existen ribosomas que sintetizan proteínas y son más pequeños que los del citoplasma de la célula. De hecho, tienen el tamaño y la composición del RNA de los ribosomas de los procariotas.
Por todas estas razones, la hipótesis del origen simbiótico de las mitocondrias y de los plastos halló adeptos en los años 1970, como Lynn Margulis, de la Universidad de Amherst (Massachusetts), y Sarah P. Gibbs, de la Universidad McGill en Montreal. La biología molecular subrayó todavía más el parentesco entre estos organelos y los procariotas. Es el caso, por ejemplo, de las zonas promotoras de los genes, es decir, de las secuencias reconocidas para proteínas que aseguran su transcripción en RNA. Se descubrieron actualmente genes homólogos (que tienen el mismo origen evolutivo) de genes bacterianos en el genoma de las mitocondrias y de los plastos. Sería sorprendente encontrar bacterias en el citoplasma de eucariotas. En realidad, semejante modo de vida no es raro entre los eucariotas: se le designa endosimbiosis. Este modo de vida es el de ciertos microorganismos patógenos, como la bacteria causante de la listerosis o de la fiebre tifoidea. Pero también, el de numerosos animales unicelulares, los protozoarios, que son colonizados por varias bacterias.
Kwang W. Jeon, de la Universidad de Tennessee, describió la aparición de una endosimbiosis entre las amebas, a raíz de una infección que diezmó un cultivo de laboratorio en 1966.
Algunas amebas sobrevivieron a la presencia de bacterias en su citoplasma: veinte años más tarde, todavía contienen unas 42 000 bacterias por célula. Pero hay más: esta asociación se ha vuelto forzosa. Las amebas ya no pueden sobrevivir si se da muerte a sus compañeros citoplasmáticos aumentando la temperatura. Otro ejemplo clásico: las plantas de la familia de las leguminosas (trébol, guisante) poseen en sus raíces unos nódulos cuyas células albergan bacterias del género Rhizobium. Estas bacterias, llegadas del suelo, ayudan a la planta a alimentarse fijando el nitrógeno atmosférico. Estos microorganismos unicelulares están rodeados siempre por dos membranas: la membrana interna, que es la de la bacteria, y la membrana externa procedente de la membrana plasmática del hospedero, que rodea a la bacteria durante su penetración en la célula. No obstante, el origen de la membrana externa de las mitocondrias y los plastos es objeto de controversia (en el caso de los plastos, contiene lípidos de tipo procariota y sería de origen mixto). ¿Cuántos sucesos de endosimbiosis han tenido lugar en el curso de la evolución? ¿Cuáles son los procariotas actuales más próximos a los que viven en endosimbiosis? La biología molecular actual puede empezar a responder estas preguntas gracias a la comparación de los genes de los organelos con los genes homólogos de los procariotas. Las secuencias de los genes se alinean y luego se comparan visualmente o por medio de programas de computadora que descubren parentescos en forma de árboles fiogenéticos. Se averigua así que las mitocondrias forman un grupo homogéneo: todas ellas descenderían de una bacteria ancestral única. Se supone que un solo suceso de endosimbiosis se produjo en un antepasado de la mayoría de los eucariotas actuales.
Se conocen descendientes directos de estos eucariotas primitivos, como el parásito intestinal Giardia. Trabajos recientes indican que el ancestro de las mitocondrias es parecido a un grupo de bacterias que usa oxígeno y que engloba, entre otros, al agente de la fiebre tifoidea, Rickettsia, y a varias bacterias simbióticas de artrópodos. Las bacterias de este grupo comparten varios caracteres con las mitocondrias, como la vida intracelular obligatoria, la utilización de oxígeno y la producción de una pequeña molécula de transferencia de energía, el ATP (trifosfato de adenosina), por medio de una vía metabólica llamada ciclo de Krebs.
Tal vez, las mitocondrias se consolidaron hace 2 000 o 3000 millones de años, en el periodo precámbrico, lo cual constituye una etapa fundamental para los eucariotas. Esta etapa supone la ventaja de la respiración celular: sin mitocondrias, la célula eucariota no puede utilizar el oxígeno para oxidar los azúcares y producir su energía. Es posible incluso que los eucariotas primitivos fueran sensibles al oxígeno, como lo son en la actualidad los eucariotas carentes de mitocondrias. Las primeras mitocondrias, al consumir el oxígeno, pudieron desempeñar el papel de protector ante las células que las albergaban: el oxígeno genera derivados tóxicos para las células que no poseen las enzimas necesarias para inactivarlos.
En la actualidad, son raros los eucariotas desprovistos de estos organelos: dicha endosimbiosis parece haber sido determinante para el éxito evolutivo de los eucariotas.
Más tarde, ciertos eucariotas que originaron los diversos linajes de vegetales habrían logrado conformar los plastos, tal vez hace alrededor de 1200 o 2 000 millones de años. La diferencia entre estos linajes son los pigmentos fotosintéticos que utilizan para captar la energía luminosa. Ya se mencionaron las algas rojas. Su equipo pigmentario recuerda el de las cianobacterias (clorofila a y ficobilisomas que les dan su color rojo). Se distinguen también los vegetales verdes, que agrupan las algas verdes y sus descendientes, los vegetales terrestres: carecen de pigmentos proteicos rojos, pero están dotados de clorofila tipo a y b. La información molecular indica que los plastos verdes y rojos habrían surgido a partir de un mismo linaje. Las cianobacterias actuales son, a menudo, simbióticas y se unen a hongos, helechos o, incluso, viven en endosimbiosis en protozoos o esponjas, por ejemplo.
Lo que puede sorprender, en cambio, es que el equipo pigmentario de los plastos verdes es muy distinto al de las cianobacterias. La pérdida de los ficobilisomas y la producción de la clorofila b se produjeron varias veces en las cianobacterias durante su evolución. En 1975, Ralph A. Lewin, de la Universidad de Yale, descubrió Prochloron, un procariota carente de ficobilisomas pero que posee clorofila b. Desde entonces se han descubierto otros organismos parecidos y se ha propuesto crear para ellos el grupo de los proclorofitos. Pero la información molecular indica que, en realidad, estos organismos aparecieron en diversas ocasiones entre las cianobacterias y forman linajes diferentes, sin ninguna relación con los plastos de los vegetales verdes. Si bien los ancestros de los plastos rojos y verdes son próximos, por no decir idénticos, parece probable que en el origen de las algas rojas y las plantas verdes hubiera dos hospederos eucariotas diferentes.
En ciertas algas, los plastos presentan la impensada particularidad de estar rodeados no de dos, sino de cuatro membranas. ¿Cómo explicarlo? Estas algas pertenecen al grupo de las heterocontas: comprende las algas pardas de las costas mexicanas, como el fucus, pero también las microscópicas diatomeas. Su plasto pardo, distinto de los plastos verdes y rojos, posee las clorofilas de tipos a y c. Entre la segunda y tercera membranas, un espacio citoplasmático contiene vesículas y microtúbulos. ¿Cómo aparecieron estos plastos especiales? Un pequeño grupo de algas unicelulares, las criptofitas, atrajo la atención: estas algas también están provistas de plastos rodeados de cuatro membranas. Pero en el espacio entre la segunda y la tercera membrana hay ribosomas de tipo eucariota y un organelo curioso, el nucleomorfo, que evoca un núcleo reducido que contiene DNA. Sarah P. Gibbs planteó la hipótesis de una segunda endosimbiosis para explicar estos plastos de cuatro membranas. Entonces, el plasto comprende, del exterior al interior, una membrana de secuestro del hospedero secundario, la membrana plasmática de un alga asimilada y luego las dos membranas del plasto de esta última.
De hecho, las algas unicelulares tienen la capacidad de desarrollarse perfectamente en el citoplasma de otros eucariotas, por ejemplo, en las células de los animales que edifican los corales y las anémonas de mar de las costas mexicanas. Las algas en cuestión, ciertas dinofiageladas, existen también en forma libre y son todavía reconocibles a la perfección. Tales estados intermedios pudieron originar asociaciones más estrechas, como en las criptofitas y en las algas heterocontas.
El DNA del nucleomorfo de las criptofitas contiene genes que corresponden a los RNA de los ribosomas que lo rodean.
El genoma del nucleomorfo está formado por tres pequeños cromosomas. Estos genes son parecidos a los de las algas rojas, y los genes ribosómicos del núcleo de la célula hospedera, la criptofita, hacen que ésta esté más emparentada con los vegetales verdes.