
Si seguimos nuestro trayecto por la vía geniculoestriada, dejamos atrás el NGL, cuyas neuronas proyectan sus axones –agrupados ahora en las llamadas radiaciones ópticas– hasta la corteza visual primaria, también llamada estriada por las estrías de sustancia blanca que presenta su cuarta capa, lugar donde terminan su viaje subcortical la mayoría de las radiaciones ópticas.
El área V1, que se corresponde con el área 17 de Brodmann y se extiende a lo largo de la cisura calcarina, es la primera área cortical que recibe información directamente del NGL, pero no es, en modo alguno, la única involucrada en la visión. De hecho, es importante tener presente que el funcionamiento visual no se restringe ni tan sólo al lóbulo occipital, puesto que la información visual también se transmite directa e indirectamente a otras áreas de los lóbulos temporales y parietales. Por ejemplo, el lóbulo parietal superior recibe señales relacionadas con el movimiento y con el reconocimiento de objetos y, a través de las radiaciones ópticas, también le llega información de la periferia y del sector inferior del campo visual.
Existen, por tanto, sistemas paralelos (algunos independientes y otros semidependientes) de envío y recepción de información entre los núcleos subcorticales, el lóbulo occipital y las áreas corticales circundantes que, a su vez, se hallan especializadas en el procesamiento de diferentes atributos del estímulo (color, movimiento, reconocimiento de objetos, etc.). Es en este sentido que pensamos que V1 puede concebirse como una estación central, desde donde se puede conectar con prácticamente todas las demás estaciones de la red. Sin embargo, no siempre es absolutamente necesario pasar por V1, dado que existen caminos alternativos que permiten pasar a otras áreas de procesamiento, incluso a las más especializadas, eludiendo dicha estación central. Valgan de ejemplo los resultados obtenidos por el equipo del profesor Zeki, según los cuáles el área V5 puede ser activada por rápidos movimientos del estímulo visual sin que se detecte paralelamente activación en V1.
Este tipo de organización del sistema visual en vías funcionales paralelas permite explicar fenómenos tan curiosos como la visión ciega y otros síntomas neuropsicológicos, como veremos más adelante.
Pese a su papel como estación central, V1 también lleva a cabo un tipo de procesamiento específico de la información visual.
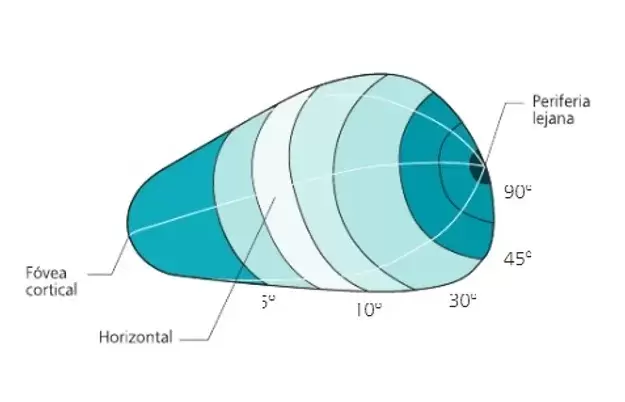
Representación de la distribución polar del mapa retinotópico que se manifi esta en V1. Obsérseve el mayor volumen que ocupan las excentricidades pequeñas, cercanas a la fóvea, y el efecto de magnificación cortical de la misma

